Начал читать «Модели…», и что-то как-то концы с концами не сходятся…
Принципиальный вопрос — каковы же всё-таки границы мезопического зрения? То есть когда включаются колбочки, и когда насыщаются палочки? Ферчайлд говорит, что «ниже 1 кд/м²» и «выше 100 кд/м²» (стр. 36). Не многовато ли? Википедия приводит другие числа: 0,01–1 кд/м² [1] или 0,03–3 кд/м² [2]. Это кажется более правдоподобным в отношении колбочек, потому что ночью цвета можно различить даже под уличным освещением, достигшем десятого этажа и отразившемся от стены. Но с другой стороны трудно поверить, что палочки отключаются настолько рано.
Вопрос принципиален в том плане, что определяет, участвуют ли вообще палочки в обиходном восприятии цвета, в том числе при просмотре отпечатков и изображений на экране монитора.
Остаётся загадкой, откуда вдруг берётся «пропорция к относительной заселённости сетчатки тремя видами колбочек» (стр. 47). Сначала на рис. 1.2 (стр. 35) приводится упрощённая схема расположения клеток сетчатки: каждой колбочке соответствует одна биполярная клетка, а той — одна ганглиозная. Потом на рис. 1.13 (стр. 48) упрощённо показывается кодирование оппонентных сигналов: каждой троице колбочек соответствует троица «арифметических узлов». Этими узлами, судя по главе «Рецептивные поля», являются ганглиозные клетки (а вовсе не биполярные, как можно было бы подумать исходя из названия; получается, что биполярные клетки служат только для объединения сигналов от нескольких палочек?). То есть вопрос в том, на какой стадии разное количество колбочек неожиданно чудесным образом объединяется в одну троицу оппонентных путей. И что происходит с избыточными L- и M-сигналами: они суммируются или усредняются?
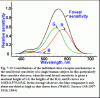
Забегая немного вперёд, уже видел рис. 8.4 (стр. 190), где эффективный диапазон колбочковых стимулов насчитывает 5 порядков. Однако на рис. 1.8 (стр. 41) и 1.15 (стр. 50) диапазон не превышает 2–3 порядка и для колбочек, и для палочек. Откуда такой разброс? Может, потому что в первом случае стимулы измеряются фотометрической яркостью, а во втором — некой «относительной энергией»?
Там же на рис. 1.15 упоминается некий «гипотетический безадаптационный ответ», по форме очень похожий на рис. 8.4, однако насчитывающий 12 порядков против 5. Что это? Никаких иных вхождений подстроки «безадаптац» и даже «гипотетич» (кроме одного единственного не по теме) во всём тексте книги найти не удалось.
Принципиальный вопрос — каковы же всё-таки границы мезопического зрения? То есть когда включаются колбочки, и когда насыщаются палочки? Ферчайлд говорит, что «ниже 1 кд/м²» и «выше 100 кд/м²» (стр. 36). Не многовато ли? Википедия приводит другие числа: 0,01–1 кд/м² [1] или 0,03–3 кд/м² [2]. Это кажется более правдоподобным в отношении колбочек, потому что ночью цвета можно различить даже под уличным освещением, достигшем десятого этажа и отразившемся от стены. Но с другой стороны трудно поверить, что палочки отключаются настолько рано.
Вопрос принципиален в том плане, что определяет, участвуют ли вообще палочки в обиходном восприятии цвета, в том числе при просмотре отпечатков и изображений на экране монитора.
Остаётся загадкой, откуда вдруг берётся «пропорция к относительной заселённости сетчатки тремя видами колбочек» (стр. 47). Сначала на рис. 1.2 (стр. 35) приводится упрощённая схема расположения клеток сетчатки: каждой колбочке соответствует одна биполярная клетка, а той — одна ганглиозная. Потом на рис. 1.13 (стр. 48) упрощённо показывается кодирование оппонентных сигналов: каждой троице колбочек соответствует троица «арифметических узлов». Этими узлами, судя по главе «Рецептивные поля», являются ганглиозные клетки (а вовсе не биполярные, как можно было бы подумать исходя из названия; получается, что биполярные клетки служат только для объединения сигналов от нескольких палочек?). То есть вопрос в том, на какой стадии разное количество колбочек неожиданно чудесным образом объединяется в одну троицу оппонентных путей. И что происходит с избыточными L- и M-сигналами: они суммируются или усредняются?
Забегая немного вперёд, уже видел рис. 8.4 (стр. 190), где эффективный диапазон колбочковых стимулов насчитывает 5 порядков. Однако на рис. 1.8 (стр. 41) и 1.15 (стр. 50) диапазон не превышает 2–3 порядка и для колбочек, и для палочек. Откуда такой разброс? Может, потому что в первом случае стимулы измеряются фотометрической яркостью, а во втором — некой «относительной энергией»?
Там же на рис. 1.15 упоминается некий «гипотетический безадаптационный ответ», по форме очень похожий на рис. 8.4, однако насчитывающий 12 порядков против 5. Что это? Никаких иных вхождений подстроки «безадаптац» и даже «гипотетич» (кроме одного единственного не по теме) во всём тексте книги найти не удалось.